Urban Evolution
Since the advent of humanity, man has been shaping the landscape around him. With the
recent global expansion of urbanised areas, man is creating a large new environment at a pace
never before witnessed in nature. In the year 2000, cities covered roughly 0.3% of the total land
area of all countries on Earth, and this number is set to increase to 1.5% by the year 2030 (1,2).
The city has its very own environmental conditions, such as climate and hydrology. One can
therefore view the city as a biome. Yes, you may not have realised it, but you are probably part
of one of the most rapidly expanding biomes on the surface of the earth! One would expect the
organisms inhabiting this relatively new urban biome to undergo evolution, since the ability to
undergo evolution is one of the most fundamental characteristics of life itself. Furthermore,
organisms are especially likely to undergo evolution when faced with new environmental
conditions, such as those they might encounter in urban habitats. The first examples of
evolution in an urban context are already starting to emerge. How did the colonisation of urban
areas influence the reproduction of a weed? How did the London Underground get its very own
species of mosquito? This article will provide you with some of the finest examples of evolution
in the latest biome to grace the surface of the earth; the urban biome.
The urban ecosystem
To understand evolution in an urban context, the unique environmental conditions that typify the
urban biome will be explored. But let us first define in general terms what makes an urban
biome 'urban'. A relatively straightforward way to classify an ecosystem as 'urban' is simply by
checking if it exhibits human population densities of at least 186 people/km2
infrastructure covers a large proportion of the land surface (3). Ecologists also like to include
less densely populated suburbs and villages that are strongly influenced by nearby urban areas,
and which themselves are urbanised to varying degrees. Only by comparing areas that exhibit
different degrees of urbanisation, can the full range of the effects of urbanisation be made clear,
as well as the thresholds of these effects (3). Now that the general definition has been clarified,
some of the peculiar and defining environmental conditions of urban ecosystems will be
discussed.
The urban heat island
Let us start with perhaps the most well-known trait of urban ecosystems: the urban heat island
effect (figure 1). This is the modification of the local climate (which generally means heating) in
urbanised areas. This is caused primarily by absorption, and trapping of, solar shortwave
radiation (heat) by non-reflective and reflective building materials, respectively. Besides
retaining heat, cities also generate it. This so-called anthropogenic heat is the result of
combustion processes inherent to industry, traffic and the heating of buildings (4). The
discrepancy between urban and nearby rural temperatures varies seasonally. In midlatitude
U.S. cities for example, temperature is increased approximately 1-2°C in the winter and
0.5-1.0°C in the summer (5). The same combustion processes that are largely responsible for
this heating also contribute to pollution of the urban atmosphere. This changes the local
atmosphere, such that it has an increased absorption and re-emission of longwave radiation,
causing a local greenhouse effect. Another effect of air pollution is that it causes increased
precipitation in cities and the area downwind of them. This is because air pollution is usually
related to higher concentrations of dust in the atmosphere, which helps water droplets form (5).
This brings us to a second peculiarity of urban ecosystems: their hydrology.
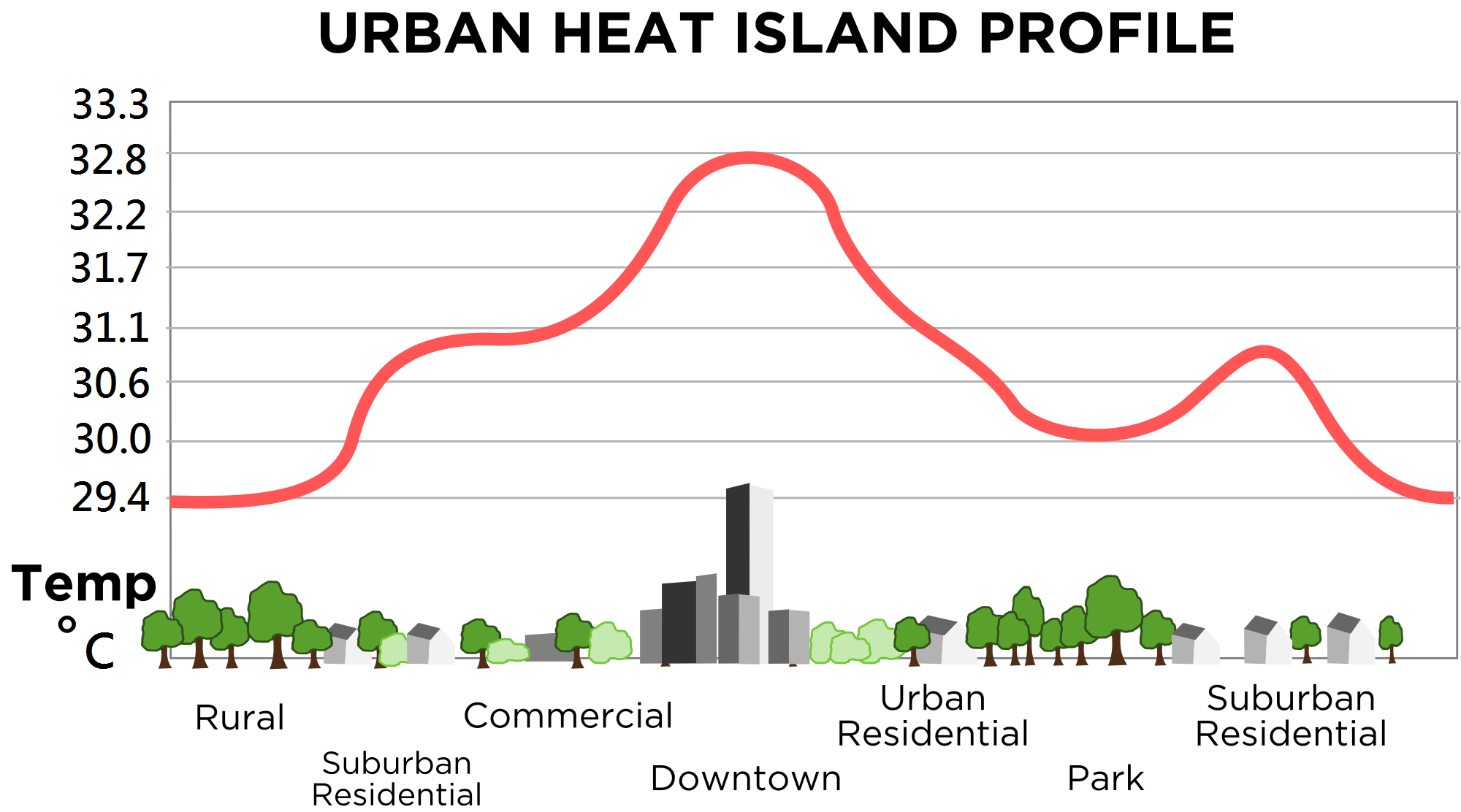
Figure 1:
The urban heat island effect along an urbanisation gradient. Source.
Hydrology
The hydrology of cities is distinct due to differences in inflow and outflow of water. Precipitation
is enhanced in, and downwind of, cities as a result of the higher concentrations of dust and air
pollution in urban atmospheres. Precipitation can be up to 5-10% higher in cities, which causes
cities to generally experience more cloudiness and fog than neighbouring non-urban areas. The
relationship between air pollution and rainfall is reflected clearly in a cyclic precipitation pattern.
Manufacturing processes and transport release particulates into the local atmosphere, which
build up during the workweek. This leads to an increased probability of precipitation at the end
of the work week (5).
In non-urbanised areas, plants and soil take up most of the precipitation. This absorption gives
water time to take up resident heat, evaporate, take the heat with it and cool down the
environment (4). A feature characteristic for urbanisation is the general reduction of
perviousness of the ground to precipitation. This has everything to do with impervious surface
cover, think of things like buildings and asphalt. Together with efficient drainage systems and a
relative lack of vegetation, this causes the water to be drained away before most of it can
evaporate. This contributes to the urban heat island and keeps groundwater levels relatively low (6).
Fragmentation
Another defining characteristic of urban ecosystems is that the habitats they contain are
generally highly fragmented. Habitat patches are isolated by roads, buildings or intensively
managed agricultural land. Thus, with increasing human population densities, the patch size of
semi-natural habitats tends to decrease. Fragmentation due to habitat loss, is bound to cause
a loss in biodiversity inside the patches, although there is more to it than that(7).
Urban biodiversity is certainly more than the number and abundance of species within a single
type of habitat. Yet another characteristic of the urban landscape is the wide variety of habitat
types. Due to this variability, the diversity in species composition in different habitat patches is
high in cities. So, despite the fact that urban green spaces are isolated and small, species
richness at the landscape level is often fairly high in urban areas(8). This diversity of patches in
cities, somewhat compensates for the loss of biodiversity due to degradation of the individual
patches and the loss of habitat.
Defining evolution
To understand evolution in an urban context, one first has to understand some concepts of
evolution. The word evolution is generally used synonymously with change. To modern day
biologists, however, it has a more refined meaning that can be phrased as 'a change in the
properties of groups of organisms over the course of generations' (9). These properties are
largely governed and carried across generations by particular pieces of DNA (or RNA) within
any organism's genome, called genes. It is the alteration of the frequencies of different variants
of genes, alleles, that is often the focus of evolutionary research (10,11,12,13,14).
Variation
The very foundation of evolution is genetic variation. This variation is primarily caused by
recombination and mutations, which arise continuously and at random (9. Genetic variation is
closely tied to phenotypic variation, and works in concert with environmental influences.
Alteration of the genetic and in particular allelic variation is the central process of evolutionary
change. There are several mechanisms that govern the alteration of allele frequencies
throughout generations, which are the actual causes of evolution. Genetic drift and natural
selection are the first and foremost of these mechanisms (9).
Genetic drift
Like mutations, which are the foundation of evolution, genetic drift is a random process. It can
best be described as the random fluctuation of allele frequencies due to 'sampling error' (9).
This basically means that a sample of alleles that passes from one generation to the next, is, by
chance, not representative of the alleles that were present in the previous generation (figure 2).
Genetic drift is best visible when looking at alleles that are equal in fitness to each other:
selectively neutral alleles. This is due to the fact that drift is essentially random and any kind of
selective influence is not. So what is fitness? Fitness is the probability that an individual with a
particular phenotype or genotype passes on its genes. This consists of the probability that the
individual survives to reproductive age, and the average number of offspring an individual has,
depending on its (if any) gender (9). Genetic drift is essentially a neutral process that will cause
selectively neutral alleles to fluctuate and even fixate in a population over the course of
generations. If it were the only factor in play in evolution, fixation would occur for all kinds of
random of alleles, regardless of their effect on fitness.
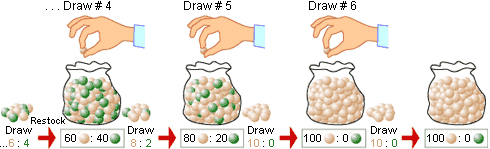
Figure 2:
Figure illustrating fixation of an allele through sampling error. If the different alleles (marbles)
pass randomly as a sample from one generation to the next they are bound not to accurately represent
the previous generation, simply by chance. The exception is if there is just one allele left, which means
that it has been fixed in the population. Source.
Natural selection
Contrary to genetic drift, natural selection is the only process within evolution that may result in
and select for adaptation. Genetic drift may also result in adaptation but does not select for it.
This is why genetic drift can be seen as one of the drivers behind non-adaptive evolution. The
definition of an adaptation is relatively straightforward. It can be defined as a characteristic that
enhances the survival or reproduction of organisms that bear it, relative to alternative character
states (9). Survival in this definition refers primarily to the chance of survival until reproductive
age, since natural selection only directly selects for adaptations that enhance reproductive
success. A trait will only be under positive selection if it makes an organism more likely to
survive longer and also increases its relative reproductive success. This is because ,eventually,
it all boils down to if and how often an individual can pass on its genes relatively to individuals
with different genotypes or phenotypes, that is to say, its relative fitness.
Something to always keep in mind is that natural selection does not have foresight and
therefore does not act purposefully. It is essentially nothing more than a name for statistical
differences in reproductive success among biological entities (9).
Adaptation to pollution
Since the urban biome originated relatively recently, with regards to evolutionary timescales, the
amount of both adaptive and nonadaptive evolution that might have already taken place, is
expected to be limited. Nonetheless, recent studies have already provided some marked
examples of evolution at work in urban environments, which will be discussed below. If one
were to compile a list of activities at which the species Homo sapiens excels, one would have to
include polluting. The urban environment, filled to the brim with humans, may often be
especially polluted. Pollution is expected to impose a rather strong selective pressure on a
population since it can cause direct mortality or impede growth. Urban soils are often
extensively polluted with heavy metals such as chromium, copper, iron, zinc and lead, that
originate from all sorts of human activities (15,16). It may therefore not come as a surprise that
many adaptations to pollution have occurred amongst disparate taxonomical groups. Some
examples include: bacteria breaking down xenobiotic organic compounds (17), fungi adapting
to high levels of radiation, such as those inside the Chernobyl nuclear reactor (18), killifish
adapting to polychlorinated biphenyl (PCB) (19) and plants adapting to metal pollution (20, 21).
Metal tolerant plants
Experiments with the liverwort Marchantia polymorpha (figure 1) have shown that urban
populations of this liverwort have a high tolerance of lead. Furthermore, specialised ecotypes
that were more resistant to copper pollution were found in some other bryophytes (20).
However, adaptation to heavy metal pollution is not just limited to bryophytes, similar tolerances
were also found in numerous flowering plants, such as Arabidopsis halleri, Allysum bertolonii
and Silene vulgaris (figure 3) (21). In the latter case, specialised ecotypes for copper, zinc or
cadmium were also identified (22). Specialised ecotypes refer to the fact that populations were
either relatively tolerant or relatively intolerant to pollution with certain metals. Such a
differentiation into two distinct ecotypes may be viewed as a precursor to the formation of a new
species, through a process known as parapatric speciation. This process which will be
explained in more detail later on.
Metal tolerant Silene vulgaris plants were found to constitutively exhibit higher levels of the
amino acid proline, even when they were not actually exposed to heavy metals. Several
different hypotheses exist regarding the role of these higher proline levels in metal tolerant
plants. First of all, proline might serve as a signalling molecule, activating the plant's response
to osmotic stress, which can be caused by the uptake of heavy metals. Furthermore, there are
actually some indications that proline plays a role in the active binding of heavy metal ions.
Finally, proline is also able to serve as an antioxidant, by scavenging the reactive oxygen
species that are often produced as a result of heavy metal uptake (21).

Figure 3: Several plants of which metal tolerant ecotypes are known.
Top-left: Marchantia polymorpha. Source.
Top-right: Silene vulgaris. Source.
Bottom-left: Allysum bertolonii. Source.
Bottom-right: Arabidopsis halleri. Source.
Crepis sancta: adaptation to fragmentation
Another plant that has successfully adapted to urban environments is Crepis sancta. This
annual weed with yellow flowers belongs to the Asteraceae family (figure 4). Like in all
members of the Asteraceae, the flowers of Crepis sancta are grouped in so-called heads:
bundles of many minute flowers grouped together. Most of the flowers in each head (80-100)
produce seeds to which fibres are attached. These fibres form a parachute-like structure,
named pappus, very similar to that found in dandelions (figure 5). The pappus allows the seeds
to be carried by the wind, thereby aiding seed dispersal. In Crepis sancta, each head also
contains some flowers (10-15) that form seeds to which no pappus is attached. Since these
seeds have a limited dispersal capacity when compared to the seeds with pappus, these seeds
are often referred to as non-dispersing seeds(23).

Figure 4: Crepis sancta, in an urban environment. Source.
The ratio between the number of non-dispersing and the number of dispersing seeds in each
head can be calculated and is known as the R-ratio. Researchers have hypothesised that this
R-ratio might be higher in urban populations of Crepis sancta, due to the highly fragmented
nature of the urban environment. In a highly fragmented environment, where suitable patches lie
far apart, the chance of the dispersing seeds landing in a patch of suitable habitat is fairly small,
because dispersing seeds are likely to end up outside the patch in which the parent plant is
located. The chance of ending up inside a new suitable patch is also minimal. The chance that
the non-dispersing seeds will end up in a suitable patch is rather high, since they will most likely
land in the very same patch in which the parent plant is located. So, as the distance between
patches of suitable habitat decreases i.e. the fragmentation increases, the chance that the
dispersing seeds will land in a suitable patch also decreases. In other words, the cost of
dispersal increases. Therefore, it should be advantageous to increase the ratio between non-
dispersing and dispersing seeds, thereby investing relatively more resources into the production
of non-dispersing seeds, that will end up in a suitable patch of habitat for sure (23).

Figure 5: A head of Crepis sancta showing several dispersing seeds with their attached pappus.
Source.
Subsequently researchers examined the R-ratio of populations of Crepis sancta that occurred in
patches surrounding trees on the sidewalk of Montpellier. This R-ratio was then compared to the
R-ratio of populations of Crepis sancta that lived in nearby, large, relatively unfragmented rural
areas. The R-ratio of the urban populations was found to be roughly 4.5% higher. Using
quantitative measurements of the dispersal chances and the heritability of the R-ratio, the
researchers were even able to estimate that this feat of rapid evolution took place over roughly
5-12 generations of selection, which was consistent with building dates of the sidewalks that
were examined (23).
Speciation
One of the most essential evolutionary processes that has not yet been discussed in an urban
context is speciation. Speciation, the process through which new species are formed, is the only
process that preserves information about the evolutionary history of lineages. Speciation is the
very reason there is such a thing as biodiversity. Assuming we only take the Biological species
concept into account, we can define speciation as the evolution of reproductive barriers. There
are four variants of this definition of speciation. First is peripatric speciation, more commonly
known as founder effect speciation, in which sampling error is at the genetic base of new
reproductive barriers. The second variant is called sympatric speciation, which is speciation
within a, at first, randomly mating population. Allopatric speciation is most easily observed
and occurs between two populations that become geographically or otherwise physically
isolated. Allopatry is not defined by geographical distance but by severe reduction of gene flow
between populations. Gene flow is limited to such an extent that very weak selection or even
genetic drift can cause divergence (9). It is not hard to imagine that this type of speciation is
likely to be prevalent in the highly fragmented urban landscape. Finally there is parapatric
speciation, a type of speciation that was already alluded to before in the context of metal
tolerant plants. In parapatric speciation, neighbouring but spatially distinct populations diverge,
despite some ongoing gene flow. Parapatric speciation can occur if gene flow between two
populations is weaker than the distinct selective pressures on different gene combinations.
Hybrids of the two population would be unable to survive in either population since they would
lack the right combination of genes to be competitive (9).
Urban cave systems and the tale of the underground mosquito
Cave systems provide fertile ground for evolution, as is illustrated by the fact that they are often
inhabited by species that only occur within a specific cave system (24). What you may,
however, not have realised is the fact that many cities have their own man-made "cave
systems". In the city of London, it is a rather extensive one at that: the London underground. It is
in this 100-year-old underground network that we find the only probable case of speciation in an
urban environment known to the authors. This is especially significant since, as mentioned
earlier, speciation is the only process that conserves information about the evolutionary history
of a certain lineage. So let us consider the curious case of Culex pipens: the common house
mosquito (figure 6).
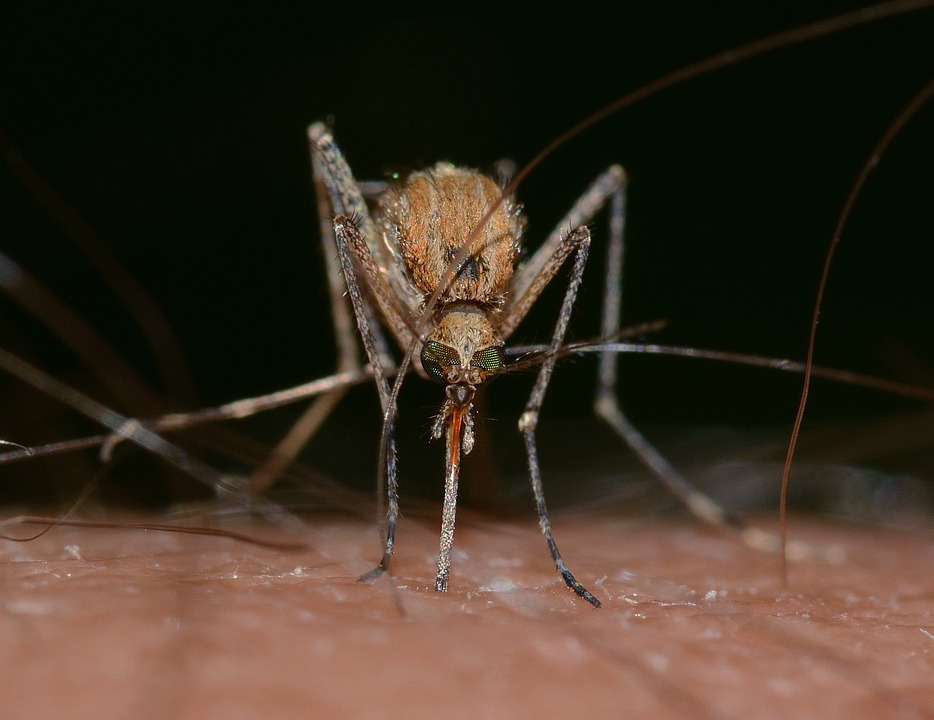
Figure 6: Culex pipens, drinking the blood of a large, relatively hairless mammal from the genus Homo.
Source.
In 1998, two biologists decided to examine the genetic differentiation between the populations of
Culex pipens that occurred aboveground and the populations that inhabited the London subway.
What made this investigation particularly interesting, was the fact that the underground
populations were known to exhibit several ecological traits that distinguished them from the
aboveground populations. The underground forms were known to be active all year round and
preferably bite mammals, whilst the aboveground forms do show a winter diapause and prefer
to bite birds. Furthermore, the females of the underground populations are able to lay eggs
without the consumption of a blood meal, whilst the females of the aboveground populations
required a blood meal before depositing their eggs. Morphologically however, the two
populations are indistinguishable (25).
What the researchers uncovered was beyond what anyone could have imagined: very little
gene-flow occurs between the two populations. The real stunner? Laboratory breeding
experiments confirmed the inability of the aboveground and underground populations to
interbreed. According to the biological species concept, these two populations should thus be
viewed as two separate species! Furthermore, the results of the genetic study indicated that the
underground species had originated from a single colonisation event, since the amount of
genetic variation within the underground population was severely reduced. This single founder
event clearly points to the occurrence of allopatric speciation. Subsequently, the researchers
found that most of the genes found in the underground populations were unique to mosquito
populations in great Britain, indicating that these mosquitoes originated from an invasion of local
aboveground Culex pipens. Since most of the London Underground is less than 100 years old,
we now have a clear cut example of rapid allopatric speciation in the urban environment. End of
story, right (25)?
The London Underground mosquito: modern insights
First of all, it was recently revealed that the underground form of Culex pipens seems to have
spread to numerous other cities as far away as Russia. A Russian study that utilised more
modern DNA-sequencing techniques, reaffirmed the results from London: the urban
underground mosquitoes seem to comprise a separate species, since no gene-flow occurs
between the underground and aboveground populations. Furthermore, this study supported a
single origin for the underground population. In 2004, a study was published that also supported
the single origin of the underground populations. This study, however, also uncovered evidence
that the underground populations in northern Europe were not derived from local populations,
but had instead originated from southern populations. In these southern populations, two
different forms of C. pipens occur, one exhibiting the ecological traits of the northern
underground populations, known as the molestus form, and another one exhibiting the
ecological traits of the northern aboveground populations, known as the pipens form. In Africa
and the Middle East these forms are not able to hybridise (26,27).
So the northern European underground species, although it should certainly be considered a
separate species, seems not to have originated from recent local adaptation to an underground
urban lifestyle. Instead, it seems to be the product of an invasion of the southern molestus form.
However, the subterranean molestus form could still provide us with the first example of urban
speciation, but only if the northern underground molestus mosquitoes have somehow become
genetically isolated from their southern molestus counterparts. Something that is not at all
unlikely due to the fact that the northward migration of the molestus form seems to have been a
one-off event, since the northern underground populations have a single origin. If subsequent
gene-flow between the southern and northern molestus populations does not occur, genetic drift
will tend to eventually bring about reproductive isolation between the northern underground
molestus population and the southern molestus population. This would provide us with a, be it
somewhat more complicated, parapatric speciation event in an urban context. To test if this
event has already occurred, interbreeding experiments between the northern underground
molestus and the southern molestus should be conducted, to test their degree of reproductive
isolation. Such experiments have, however, not yet been performed. Therefore the first truly
conclusive evidence of urban speciation remains to be uncovered (26,27).
The prize of the first truly urban species may, however, not be awarded to the London
Underground mosquitoes. Modern molecular techniques are making it easier than ever before
to detect genetic differentiation between urban and rural populations. Recent molecular
investigations have indicated the occurrence of rapid adaptive evolution in urban white-footed
mice living in New York (28). Allowing for the suggestion that these mice may eventually
become genetically isolated from their rural counterparts.
Conclusion
Urban areas can be considered a true biome due to their own unique ecological characteristics.
As expected, we have uncovered some of the earliest examples of urban evolution. Species
have adapted to both pollution and fragmentation whilst conquering the urban biome. The first,
somewhat disputable, case of urban speciation has also been illustrated. One must, however,
realise that the urban biome is still relatively young and that more examples of urban evolution
are bound to be uncovered. The detection of the first urban speciation event is merely a matter
of time, if the London Underground mosquitoes or the white-footed mice of New York fail to
provide it, some other species will. The authors would therefore like to encourage future
biologists to study and document the emerging world of urban evolution.